自閉症譜系障礙(Autism Spectrum Disorder,簡稱 ASD)正在改寫我們對人類大腦多樣性的理解。從 2000 年美國每 150 名兒童中有 1 人確診,到 2022 年急升至每 31 名兒童中就有 1 人——這個數字背後不僅是診斷意識的提升,更是一場科學認識的革命。全球範圍內,世界衛生組織估算約每 127 名兒童中就有 1 人處於自閉症譜系,影響著超過 7,800 萬人。
然而,自閉症仍然是醫學科學中最複雜的謎題之一。它不是一個單一的疾病,而是一個涵蓋極其廣泛表現的譜系——從需要全天候支持的深度障礙到高功能自閉症患者。這種異質性使得「自閉症的成因」這個問題本身就帶有誤導性:我們可能面對的不是一個問題,而是數百個問題交織在一起的網絡。過去五年,基因組學、神經影像學和計算神經科學的突破正在以前所未有的速度拆解這個網絡。從 CRISPR 基因編輯猴子模型到萬人規模的隊列研究,從腸道微生物組到人工智慧輔助早期診斷——自閉症研究正在經歷一場真正的範式轉移。
本文將系統性地梳理自閉症成因與治療的最新科學圖景,揭示這場從「症狀管理」到「機制理解」的深層轉變。
在理解自閉症之前,我們必須先面對一個根本性的流行病學事實。自閉症的「患病率」在過去二十年間出現了戲劇性的增長。2026 年,全球疾病負擔研究(GBD 2023 Collaborators)在《柳葉刀》上發表了覆蓋 204 個國家和地區的系統性分析,確認了這一持續上升的趨勢。
美國疾病控制與預防中心(CDC)的數據最為完整。2000 年,美國自閉症患病率為每 150 名兒童中 1 人(0.67%);2010 年上升至每 68 人中 1 人(1.47%);2016 年為每 54 人中 1 人(1.85%);而 2022 年的最新數據已達到每 31 人中 1 人(約 3.2%)。這個增長速度遠遠超出了單純的診斷標準變化所能解釋的範圍。
專家共識認為,患病率的上升是多重因素疊加的結果。1994 年 DSM-IV 的診斷標準擴大了自閉症的定義範圍,將亞斯伯格症納入譜系;2013 年 DSM-5 進一步用「自閉症譜系障礙」統一分類,消除了亞型之間的界限。與此同時,公眾意識的提升、早期篩查的普及、服務可及性的改善,以及學校和醫療系統中診斷路徑的完善,都推動了更多個案被識別。但流行病學家也承認,真正的患病率上升——即環境因素的貢獻——無法被完全排除。
性別比例是一個備受關注的現象。男性被診斷的比率約為女性的三倍。Cruz 等人(2025 年)在《 Neuropsychology Review》上發表的系統性回顧和薈萃分析確認了這一比例,但同時指出診斷中可能存在男性偏差。Maciver 等人(2026 年)在《 Autism》期刊上發表的追蹤研究令人鼓舞地發現,我們正在變得越來越善於識別神經多樣性的女性和女孩。自閉症女性傾向於表現出較少的非典型行為,因此可能被低估或診斷延遲——這被稱為「女性保護效應」的假說。
共病症同樣值得關注。約 25-32% 的自閉症患者同時患有 ADHD;30-40% 伴有智力障礙;約 10% 患有癲癇;焦慮症(17-23%)和抑鬱症(9-13%)的共病率也顯著高於普通人群。睡眠障礙和胃腸道問題在自閉症群體中極為常見。另一個近年的發現是:自閉症群體中 LGBTQ+ 身份認同的比例高於一般人群,Pereira 等人(2026 年)在《 Autism》上報告了跨性別和性別多樣化的自閉症青少年面臨更高的抑鬱風險。
如果自閉症研究有一個最核心的結論,那就是:遺傳因素主導了自閉症的風險。雙生子研究一貫顯示遺傳率在 60% 至 90% 之間。如果一個家庭中已經有一個自閉症孩子,第二個孩子的復發風險為 7% 至 20%——遠高於普通人群的基線風險。
但「遺傳」絕不意味著「單一基因」。目前已知與自閉症相關的基因超過一百個,但其中絕大多數只解釋了不到 1% 的案例。這是一個典型的複雜遺傳性疾病——數百個基因各自貢獻微小效應,通過多基因風險評分(Polygenic Risk Score)的累積效應決定最終的風險。
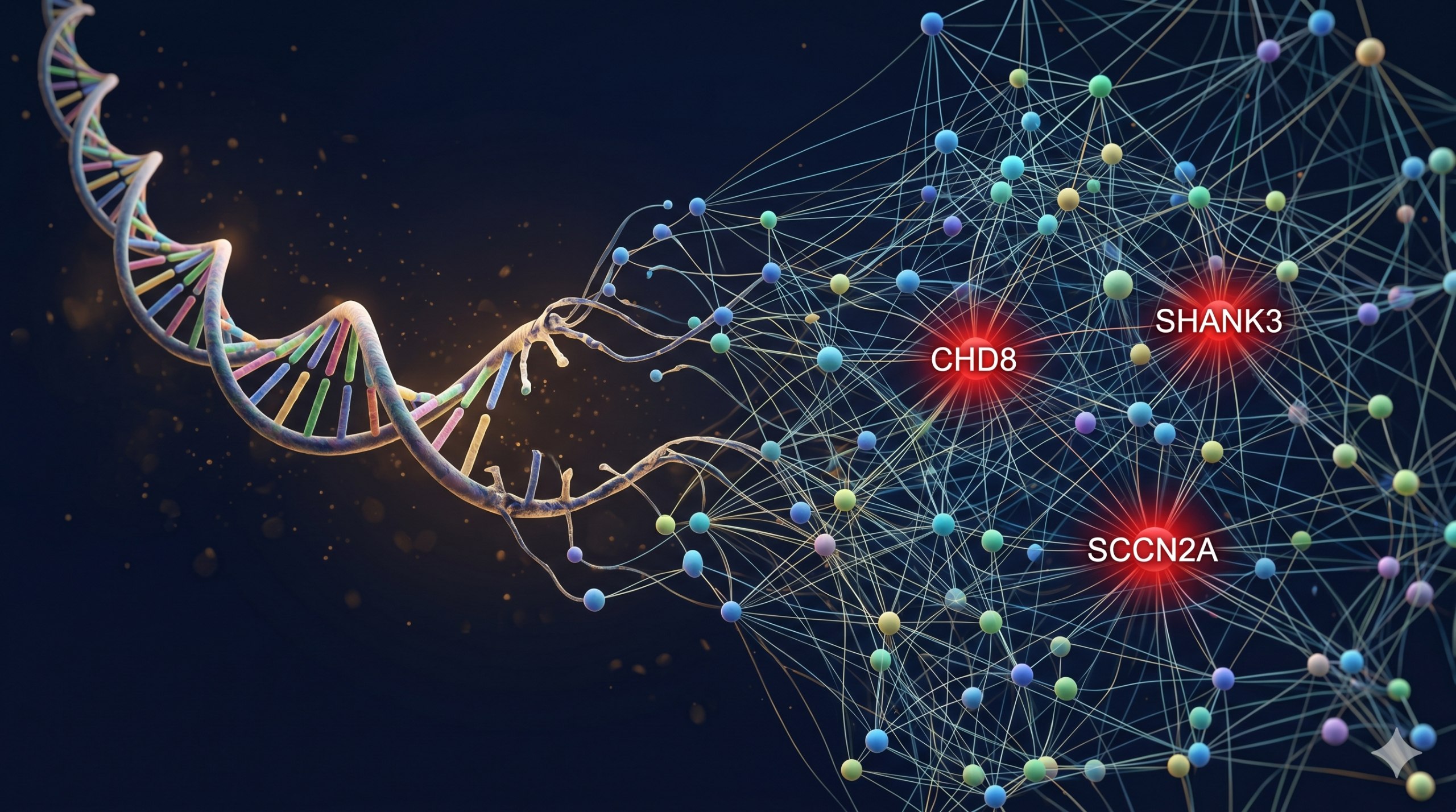
在已知的基因中,有幾個尤其值得關注。CHD8 是自閉症中最常見的 de novo(新發)突變基因。Nitahara 等人(2026 年)在《自然·通訊》上發表的研究揭示,胎兒中期的 Chd8 突變會導致腹側神經發生缺陷,從而在小鼠中驅動自閉症樣行為。SHANK3 編碼突觸後支架蛋白,其突變導致 Phelan-McDermid 症候群——一個嚴重的綜合症型自閉症。2026 年,Jiang 等人在《神經元》上發表了里程碑式的研究:利用 CRISPR 技術創建了首個 SHANK3 突變的獼猴模型,展示了行為表型和神經元生物標誌物。SCN2A 編碼鈉離子通道 Nav1.2,其功能喪失型突變同時與自閉症和癲癇相關。SYNGAP1 編碼突觸 Ras GTP 酶激活蛋白,其相關疾病的精準治療正在成為基因治療的重要方向。
約 30-40% 的自閉症案例涉及 de novo 突變——即在配子形成或胚胎發育早期新發生的突變,並非遺傳自父母。這些突變在伴有智力障礙的自閉症個體中更為常見。突變率隨父親年齡增長而增加,為高齡父親與自閉症風險之間的流行病學關聯提供了生物學解釋。
除了單基因變異,拷貝數變異(Copy Number Variations, CNVs)也扮演著重要角色。16p11.2 的缺失或重複是最常見的 CNV 之一,而 22q11.2 缺失(DiGeorge 症候群)與自閉症風險升高密切相關。綜合症型自閉症約佔所有案例的 25%,包括脆性 X 症候群、結節性硬化症、Rett 症候群和 Angelman 症候群等已知遺傳疾病。
值得注意的是,遺傳並非靜態的宿命。表觀遺傳學揭示了環境因素如何通過改變基因表達而不改變 DNA 序列來影響自閉症風險。Han 等人(2025 年)在《生物學》期刊上綜述了早期生活壓力與自閉症在下丘腦-垂體-腎上腺軸關鍵基因表觀遺傳學上的匯合。Ebenezer 等人(2026 年)在《細胞》上報告,母體炎症會改變新生兒大腦單核細胞中的核和線粒體 DNA 甲基化模式。這些發現正在搭建遺傳與環境之間的因果橋樑。
遺傳雖然主導風險,但環境因素在自閉症的發病機制中同樣不可忽視——尤其是在特定的遺傳易感性背景下。孕期因素是研究最集中的領域。母親在孕期感染風疹已被確認會顯著增加自閉症風險。更廣泛地說,母體免疫激活(Maternal Immune Activation, MIA)是確立的風險因素。Spagnuolo 等人(2026 年)在《大腦、行為與免疫》上發表的研究提供了令人振奮的發現:早期阻斷 IL-17A 信號可以預防 MIA 引起的行為異常和海馬突觸變化。Gargus 等人(2026 年)進一步探索了迷走神經刺激作為抗炎療法來逆轉 MIA 誘導的子代小膠質細胞改變,展示了從機制理解到干預的潛在路徑。
藥物暴露同樣受到關注。丙戊酸(一種抗癲癇藥物)是已確立的致畸劑,孕期使用會顯著增加自閉症風險。對乙酰氨基酚(撲熱息痛)的潛在風險仍在爭議中。Cramer 等人(2025 年)在《產科與婦科學》上的研究和 Balkanas 等人(2026 年)在《自閉症與發育障礙雜誌》上的討論都指向這一方向,但因果關係尚未明確建立。葉酸的補充則呈現出更複雜的圖景:Vasconcelos 等人(2025 年)和 Abate 等人(2025 年)的研究發現,孕期葉酸缺乏和過量均可能與自閉症風險相關——劑量的 U 型曲線是公共衛生乾預的重要提示。
空氣污染是另一個持續積累證據的領域。Kang 等人(2026 年)利用新生兒代謝組學追蹤產前空氣污染暴露與自閉症風險之間的分子連接。Ayoub 等人(2026 年)在《兒童心理學與精神病學雜誌》上發表了基於全國人口隊列的研究,確立了產前環境空氣污染暴露與神經發育障礙風險之間的相關性。這些研究的樣本量從數萬到數十萬不等,為公共衛生政策提供了堅實的流行病學基礎。
腸道微生物組是近年興起的研究熱點。腸-腦軸異常被認為可能促發自閉症症狀。Zhong 等人(2026 年)在《大腦、行為與免疫》上發表的研究發現,羅伊氏乳桿菌(Limosilactobacillus reuteri)可以通過重新編程脂質代謝來改善 MIA 誘導的自閉症樣行為。Wu 等人(2026 年)通過整合多矩陣代謝組學揭示了腸道微生物驅動的全身代謝改變。這些發現正在將自閉症從純粹的「腦疾病」重新定義為涉及免疫系統、胃腸道和代謝系統的全身性狀況。

在科學史上,某些錯誤理論的糾正同樣重要。麻疹-腮腺炎-風疹(MMR)疫苗與自閉症之間的關聯說——源自 Andrew Wakefield 1998 年的欺詐性研究,已被徹底否定和撤稿。硫柳汞(疫苗防腐劑)也已被從疫苗中去除,但自閉症患病率並未因此下降。「冰箱母親」假說——認為冷漠的母親導致孩子自閉症——早已被揭穿。這些歷史教訓提醒我們:在神經發育科學中,簡單的因果關係往往是最不可靠的。


自閉症的大腦既非「受損」也非「異常」——它是在發育軌跡上走了不同道路的器官。神經生物學研究正在逐步揭示這條不同道路的具體特徵。
約 15-20% 的自閉症患者表現出巨頭畸形(macrocephaly),即頭圍大於正常範圍。研究顯示,自閉症兒童的大腦在早期經歷了一個過度生長階段——額葉和顳葉體積增大,而頂葉和枕葉體積正常;小腦蚓部、胼胝體和基底節則相對較小。到兒童中期,腦容量趨向正常化,但區域性的結構差異持續存在。邊緣系統的細胞更小但更密集,可能解釋了社交障礙的神經基礎。小腦中的浦肯野神經元數量和體積均減少,而小腦在情緒和語言處理中的作用正在被重新認識。
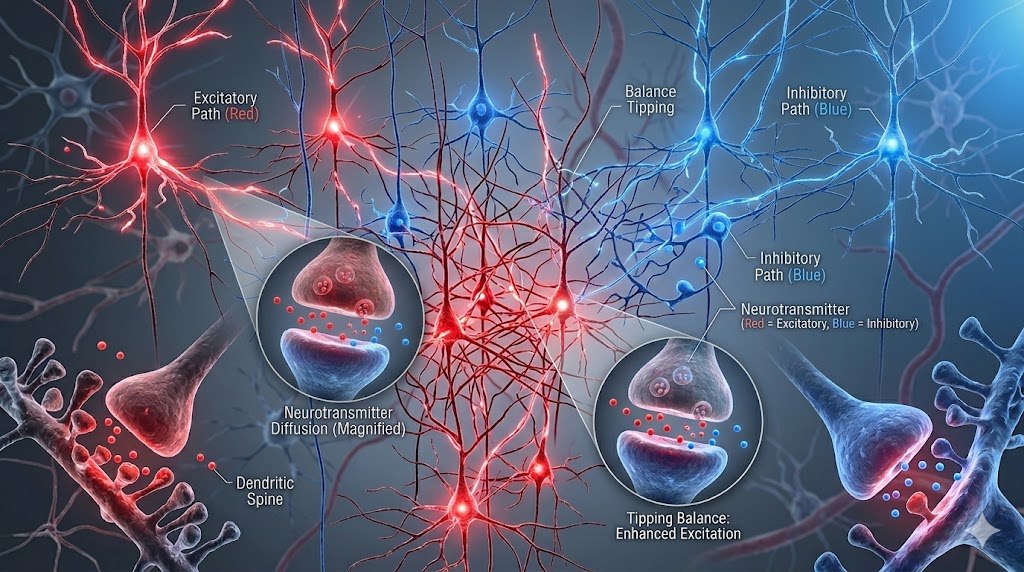
興奮/抑制(E/I)平衡假說是自閉症神經生物學的領先理論。Madia 等人(2025 年)在《AIMS 神經科學》上發表了整合遺傳學、神經遞質和計算模型的系統性綜述,為 E/I 失衡提供了多層次證據。自閉症大腦中,GABA 相關基因表達降低,穀氨酸信號改變。GABA 能抑制性傳導的減弱和穀氨酸能興奮性信號的增強,導致神經網絡處於過度興奮狀態,破壞了正常的信息處理。這就是為何像布美他尼(bumetanide)這樣的藥物——它通過阻斷 NKCC1 離子通道來恢復 E/I 平衡——成為了備受關注的治療靶點。
神經遞質系統的不平衡還延伸到其他關鍵分子。約 30% 的自閉症患者表現出血液血清素水平升高(高血清素血症),這是一個持續了數十年的神秘發現。多巴胺系統在某些自閉症亞型中發生改變。而催產素——這種與社會連結密切相關的神經肽——的研究正在從「催產素缺乏」的簡單敘述轉向更精確的理解。Boulton 等人(2026 年在《神經科學與生物行為評論》)提出了一個精準醫學框架,整合了來自動物研究、生物標誌物、遺傳學、表觀遺傳學和神經影像學的證據,解釋了為何催產素臨床試驗結果不一致:並非所有人的催產素系統都以相同方式運作。
神經炎症是另一個令人振奮的研究前沿。自閉症死後大腦組織中星形膠質細胞和小膠質細胞的數量增加,Meng 等人(2024 年)發現了小膠質細胞功能障礙。更高表達的膠質細胞和免疫細胞相關基因提示,免疫系統與神經系統之間的相互作用在自閉症發病機制中扮演著比之前認知更重要的角色。mTOR 信號通路——調控細胞生長和存活的關鍵樞紐——也與自閉症相關,通過 PTEN 和 TSC1/TSC2 等基因與自閉症風險聯繫在一起。
早期干預的效果取決於早期檢測。在過去的五年中,自閉症的早期識別技術取得了顯著進展。
M-CHAT(Modified Checklist for Autism in Toddlers)仍是最廣泛使用的篩查工具。Harper 等人(2026 年)發表了診斷準確性的複制研究;Bacopoulou 等人(2026 年)在希臘人群中驗證了 M-CHAT-R/F;Lassebro 等人(2026 年)在《JAMA 網絡開放》上將其應用於新生兒高風險人群。Chang 等人(2026 年)發現,神經發育量表的早期評估與 24 個月時的 M-CHAT 陽性篩查結果相關。
但更具變革意義的是人工智慧在診斷中的應用。Yankowitz 等人(2026 年)在《分子自閉症》上發表了基於 AI 的人際協調測量方法。V MP 等人(2026 年)在《科學報告》上展示了利用機器學習模型從 MRI 數據中識別自閉症。Alyasseri 等人(2026 年)提出了混合 EEG 特徵融合框架,使用集成學習實現準確的自閉症診斷。Xu 等人(2026 年)的 BrainPrompt+ 模型展示了從腦影像中識別神經疾病的通用方法。
眼球追蹤技術顯示,自閉症嬰兒的凝視模式在生命早期就可以被檢測到——他們傾向於較少注視他人的眼睛。Yin 等人(2026 年)在《自閉症研究》上展示了利用聲學特徵和機器學習在嬰兒中進行早期預測的可行性。Motta 等人(2026 年)發現的胎兒 MRI 生物標誌物甚至將檢測窗口推到了出生前。這些技術匯聚在一起,正在將自閉症的診斷年齡從傳統的 3-4 歲逐步前移到 12-18 個月,為早期干預創造了前所未有的機會窗口。

在行為干預領域,過去四十年的研究已經建立了一個多層次的治療譜系。
應用行為分析(Applied Behavior Analysis, ABA)仍然是證據基礎最雄厚的干預方法。早期密集 ABA(每週 25-40 小時)已被證明可以改善學齡前兒童的語言、適應功能和智力表現。Levato 等人(2025 年)發表了模塊化行為干預的隨機對照試驗結果;Anderson 等人(2024 年)比較了模塊化行為干預與綜合行為干預的效果。然而,ABA 也面臨倫理爭議——批評者指出其強調服從、可能導致提示依賴,以及利益衝突未充分報告等問題。現代 ABA 正在從嚴格的離散試驗訓練(Discrete Trial Training)轉向更自然的干預方法。
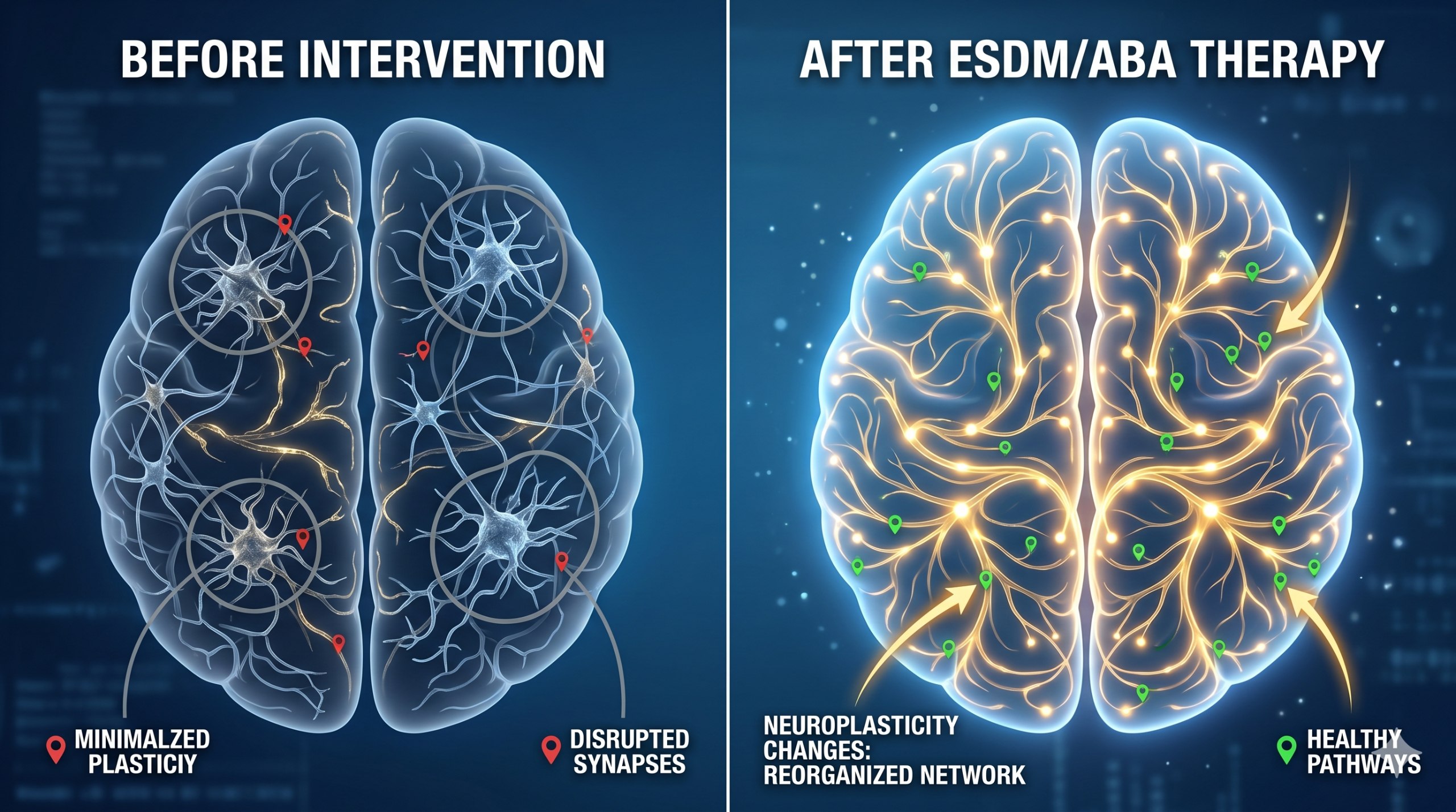
早期開始丹佛模式(Early Start Denver Model, ESDM)是 ABA 與發展原則的結合,代表了自然發展行為干預(NDBI)的主流方向。Ruta 等人(2026 年)追蹤了不同風格和強度的 ESDM 社區干預下的早期發展軌跡。Tateno 等人(2026 年)在日本驗證了 ESDM 的有效性。Yan 等人(2026 年)在《歐洲醫學研究雜誌》上揭示了 NDBI 的神經可塑性機制——這些干預不僅改變行為,還改變大腦的連接模式。
關鍵反應訓練(Pivotal Response Treatment, PRT)針對「樞紐性」領域——動機、對多重線索的反應能力、自我管理和社交發起。Schuck 等人(2026 年)整合了四項隨機對照試驗的數據,證明 PRT 改善了自閉症兒童的生活質量。Cheong 等人(2026 年)展示了遠程醫療傳遞的 PRT 的可行性——在疫情後時代具有重要意義。DIR/Floortime 關係模型強調通過兒童主導的遊戲促進情感和關係發展。TEACCH 結構化教學方法注重視覺支持和環境組織。
在藥物治療方面,進展雖然不如行為干預那樣顯著,但正在從「症狀壓制」轉向「機制靶向」。
目前,只有兩種藥物獲得 FDA 批准用於自閉症相關症狀,且它們都不針對核心症狀。利培酮(Risperidone)和阿立哌唑(Aripiprazole)被批准用於治療 5-16 歲(利培酮)和 6-17 歲(阿立哌唑)自閉症兒童的易怒、攻擊和自傷行為。兩者都是非典型抗精神病藥物,具有顯著的副作用——體重增加、代謝症候群和錐體外系症狀。薈萃分析發現,抗精神病藥物或 SSRI 在減少限制性/重複性行為方面並無顯著療效。超過 50% 的美國自閉症兒童被處方精神活性藥物或抗驚厥藥物,這反映了症狀管理的巨大需求。
在研靶點中最受關注的是催產素。Boulton 等人(2026 年)的精準醫學框架可能解釋了為何鼻內催產素的臨床試驗結果不一致——它可能只對基於特定遺傳或催產素系統變異的亞組有益。布美他尼最初是作為利尿劑開發的,但因其調節 E/I 平衡的機制而被重新用於自閉症。McNamara 等人(2026 年)的系統性綜述評估了布美他尼在神經發育障礙中的潛力,多項 II/III 期試驗結果不一。大麻二酚(CBD)同樣受到關注。Lawson 等人(2026 年)發表了 Epidiolex 在自閉症兒童中的 II 期開放標籤試驗(N=23),初步證據顯示 CBD 可能改善自閉症相關挑戰。
Balovaptan(加壓素 V1a 拮抗劑)在 II 期試驗中表現出希望但 III 期結果不確定。Arbaclofen(GABA-B 激動劑)的試驗結果同樣不一。胰島素樣生長因子 1(IGF-1)正在被研究用於 Rett 症候群和 Phelan-McDermid 症候群。
真正可能改寫遊戲規則的是新興治療方法。儘管這些方法大多仍處於臨床前或早期臨床階段,但它們代表著從「症狀管理」到「病因治療」的根本轉變。
基因治療在單基因形式的自閉症中取得了初步進展。Shokoohi 等人(2026 年)在《發現·心理健康》上系統性綜述了基因治療在自閉症中的新興角色。Roh 等人(2026 年)在《自然·通訊》上發表了一項極具說服力的研究:一種調控甘氨酸的 Slc6a20a 反義寡核苷酸(ASO)能夠恢復 SHANK2 和 SHANK3 突變小鼠及皮層類器官中的 NMDA 受體功能。這是針對自閉症核心突觸功能缺陷的分子層面干預——距離臨床應用可能仍需數年,但其概念驗證的意義不可低估。
CRISPR/Cas9 基因編輯主要用於構建疾病模型。Jiang 等人(2026 年)在《神經元》上發表的 SHANK3 突變獼猴模型是迄今最接近人類自閉症的大動物模型。Karagyaur 等人(2026 年)利用 CRISPR 創建了 Plau 基因 D277N 突變小鼠系,展示了自閉症樣特徵。這些模型不僅幫助理解疾病機制,也為未來的基因治療提供了測試平台。
神經調控正在成為一種非侵入性的治療選擇。Kang 等人(2026 年)在《腦拓撲學》上報告了一項包含 32 名兒童的隨機對照試驗:重複經顱磁刺激(rTMS)調節了自閉症兒童的靜態和動態大腦功能網絡。Tung 等人(2026 年)在《腦發育》上發表了經顱直流電刺激(tDCS)對自閉症行為症狀的系統性回顧。EEG 神經反饋訓練——教導患者自我調節大腦活動——也顯示出改善注意力和社交功能的初步證據,但研究質量限制了結論的強度。
微生物組干預是另一個令人著迷但尚處於實驗階段的方向。Peta Martinez 等人(2026 年)和 Wu 等人(2026 年)的研究分別在果蠅和小鼠模型中展示了微生物組干預挽救社交行為的潛力。糞便微生物移植(FMT)在小型試驗中顯示了改善胃腸道和行為症狀的跡象,但長期安全性數據仍然有限。

科技在自閉症干預中的角色正在快速擴大。McCleery 等人(2026 年)在《自閉症與發育障礙雜誌》上發表了一項隨機對照試驗:虛擬現實(VR)干預可以幫助自閉症患者學習如何與警察互動。Cui 等人(2026 年)的薈萃分析證實了 VR 運動對自閉症兒童社交技能和情緒識別的積極效果。可穿戴設備也在被用於生理監測(心率、皮膚電導)、睡眠追蹤和活動監測。Lee 等人(2026 年)綜述了可穿戴技術在兒童神經發育障礙中的應用。Bergwerff 等人(2026 年)則為可穿戴設備在兒童青少年心理健康中的使用提供了範圍綜述。
從全面的科學審視中,可以提煉出幾個核心洞察。
第一,自閉症正在從「行為定義的症候群」轉向「生物學定義的疾病集合」。遺傳學已經揭示了數十個具有不同機制的高置信度風險基因。隨著基因分型的普及和成本下降,自閉症的分子亞型化將成為現實。這意味著「自閉症」這個術語在未來可能更像「癌症」——一個涵蓋數百種不同疾病的統稱,每種疾病都需要針對性的治療策略。
第二,E/I 平衡理論為理解自閉症的神經生物學提供了一個統一框架。從基因突變(SHANK3、SCN2A)到神經遞質異常(GABA、穀氨酸),再到藥物靶點(布美他尼、Arbaclofen),E/I 失衡就像一條貫穿線將分離的觀察連接起來。這一框架的最大價值在於它提供了可量測的生物標誌物和可干預的治療靶點。
第三,早期干預的窗口正在被不斷前移。AI 輔助診斷、眼球追蹤、聲學分析和胎兒 MRI 都在將檢測年齡推向更早的階段。結合 ESDM 等自然發展行為干預,我們可能能夠在神經可塑性的關鍵期內更有效地改變發育軌跡。但這也帶來了倫理問題——何時進行早期篩查是適當的?標籤化的風險如何管理?
第四,藥物治療正在從神經精神藥物轉向機制靶向治療。催產素、布美他尼和 CBD 代表了第一波「機制藥物」,而基因治療和 ASO 則代表第二波。但在這個過渡期,管理期望至關重要——從實驗室到臨床的距離仍然遙遠。
最後,自閉症研究正在經歷一場根本性的認識轉變:從「尋找治癒」轉向「理解與支持」。神經多樣性運動強調自閉症不是一種需要修復的缺陷,而是一種人類認知的多樣化形式。這一觀點與科學研究並不矛盾——更好的理解帶來更好的支持,無論目標是改善生活質量還是發展新的治療選項。真正重要的不是消除自閉症,而是消除自閉症患者在生活中面臨的障礙——從教育系統的包容性到就業市場的接納,從醫療服務的可及性到社會態度的轉變。
對全球約 7,800 萬自閉症患者及其家庭來說,這些科學進展正逐步轉化為更早的識別、更有效的干預和更有尊嚴的生活。
成年自閉症:被遺忘的一群
在自閉症研究中,一個長期被忽視的群體是成年患者。兒童自閉症的研究獲得了絕大部分資源和關注,但自閉症是終身狀況。Lage 等人(2026 年)在《巴西精神病學趨勢》上探討了成人自閉症的臨床建構和歷史洞見。Garza Guerra 等人(2026 年)在《Cureus》上指出了成人精神科實踐中對自閉症的低識別及其影響。Williams 等人(2026 年)提出的 OET 自閉症框架重新思考了青少年和成年女性自閉症患者的臨床實踐。Mair 等人(2026 年)在《自閉症》期刊上發表了一項社群媒體研究,探討神經多樣性晚期識別中的悲傷、解脫和信念轉變——這些情感的複雜交織揭示了遲來診斷的心理影響。
社群監測研究顯示,如果正式評估,相當比例的成年人會符合自閉症診斷標準。許多成年人——尤其是女性和支持需求較低者——仍然未被診斷。晚期診斷正日益被認識為一個重要的臨床問題。這不僅關乎個人身份認同的重構,更關乎獲得適當支持服務的權利。
全球差異:診斷的雙重標準
自閉症診斷的全球差異是一個被低估的問題。在低收入和中等收入國家,自閉症可能被嚴重低估。文化差異影響著症狀識別和求助行為——某些社會中正常的眼神接觸規範可能與西方標準截然不同。Ricci 等人(2026 年)在加拿大馬尼托巴省進行的人口研究探討了自閉症與移民背景的關係。Karki 等人(2025 年)報告了尼泊爾精神衛生診所的臨床社會人口學特徵,展示了截然不同的診斷圖景。
日本的普及篩查(18 和 24 個月)和英國的針對性篩查反映了不同衛生體系對早期識別的不同策略。美國對 3 歲前篩查尚未達成共識。這些差異不僅反映了資源分配的不同,更反映了對自閉症本身的文化理解差異——在某些文化中,自閉症特質可能被理解為性格特徵而非醫學狀況。
科技賦能的未來
數位療法正在崛起。Lee 等人(2025 年)開發了針對自閉症青少年的社交溝通干預手機應用。Aydemir 等人(2025 年)探索了 ChatGPT 傳遞的身體活動干預對自閉症兒童的可行性——AI 對話代理作為干預傳遞渠道的潛力令人期待。貓咪亞等人(2024 年)系統性評估了美國數位心理健康技術的應用。
虛擬現實正在從實驗工具變成治療平台。除了前面提到的警察互動訓練(McCleery 等人,2026 年),Maddalon 等人(2026 年)研究了自適應 VR 干預對自閉症兒童社交認知技能的影響。Alsem 等人(2026 年)在特殊教育環境中開發了個性化的單次 VR 社交技能訓練。Ślósarz 等人(2026 年)進一步將 VR 技術擴展到跨多種精神疾病的認知行為治療中。
可穿戴設備正在實現「現實世界中的實時監測」。Tovin 等人(2026 年)開發了用於自閉症身體活動監測的可穿戴設備依從性測量方法。Sullivan 等人(2026 年)利用可穿戴技術評估治療師在處理挑戰行為時的皮膚電導活動——這不僅是患者監測,還是治療師訓練的工具。S AA 等人(2026 年)開發了基於超圖注意力神經網絡的節能 EEG 自閉症檢測方法,為在資源有限的環境中進行低成本篩查開闢了可能。
Observatory 分析:未來的五個關鍵趨勢
站在 2026 年回望,自閉症研究正處於一個罕見的轉折點——多條獨立的研究路線正在匯聚成一個更完整的圖景。以下是 Observatory 識別的五個關鍵趨勢:
第一,生物學分型將取代行為分類。 DSM-5 的自閉症譜系概念在臨床上有用,但在科學上已經成為限制。擁有相同「自閉症」診斷的兩個人可能在遺傳學、神經生物學和治療反應上完全不同。未來五年,我們將看到基於遺傳標誌物、神經影像特徵和生物標誌物的分子亞型分類體系逐步建立。這將從根本上改變臨床試驗的設計——不再測試「自閉症藥物」,而是測試「針對 SHANK3 突變亞型的藥物」。
第二,精準醫學將從口號變為實踐。 催產素研究就是一個典範:從「給所有人催產素」到「根據催產素受體基因型和內源性催產素水平選擇治療對象」。Boulton 等人(2026 年)的精準醫學框架代表了這種轉向。同樣的邏輯將應用於布美他尼(誰的 NKCC1/KCC2 比值異常?)、CBD(誰的內源性大麻素系統失調?)和經顱磁刺激(誰的 E/I 失衡可被 TMS 校正?)。
第三,AI 將重塑診斷和干預。 從 MRI 和 EEG 的機器學習分析到大語言模型輔助的行為評估,AI 正在將診斷的客觀性和可擴展性提升到新水平。Yankowitz 等人(2026 年)的人際協調 AI 測量只是一個開始。未來,常規兒科檢查可能包含自動化、AI 驅動的發育篩查,無需專門訓練即可在基層醫療中實施。這對縮小全球診斷差距具有巨大潛力。
第四,腸-腦軸是未來五年的最大變量。 微生物組干預的安全性和可及性使其成為大規模應用的理想候選。如果 Zhong 等人(2026 年)的羅伊氏乳桿菌研究在人類試驗中得到複製,我們可能看到益生菌作為自閉症輔助治療的廣泛應用。但微生物組研究的可複制性問題——困擾整個領域的挑戰——需要嚴肅對待。
第五,自閉症話語正在從「疾病模型」轉向「神經多樣性模型」。 這不僅是社會運動,它正在重塑研究優先級和臨床實踐。治療目標從「消除自閉症」轉向「減少障礙和支持 flourishing」。這意味著不再問「如何讓自閉症患者變得更正常」,而是問「如何調整環境讓自閉症患者能夠充分發揮潛力」。這一轉變與精準醫學並不衝突——兩者都要求我們更精確地理解個體差異,無論是遺傳上的還是環境上的。

從更長的時間尺度看,自閉症研究的最終貢獻可能遠遠超出自閉症本身。研究方法——從多基因風險評分到腸-腦軸分析,從早期干擾的神經可塑性到神經多樣性的社會模型——正在為理解整個人類大腦的發育和功能提供新的框架。自閉症不僅是人類神經多樣性的一種表現,它正在成為解開大腦秘密的一把鑰匙。